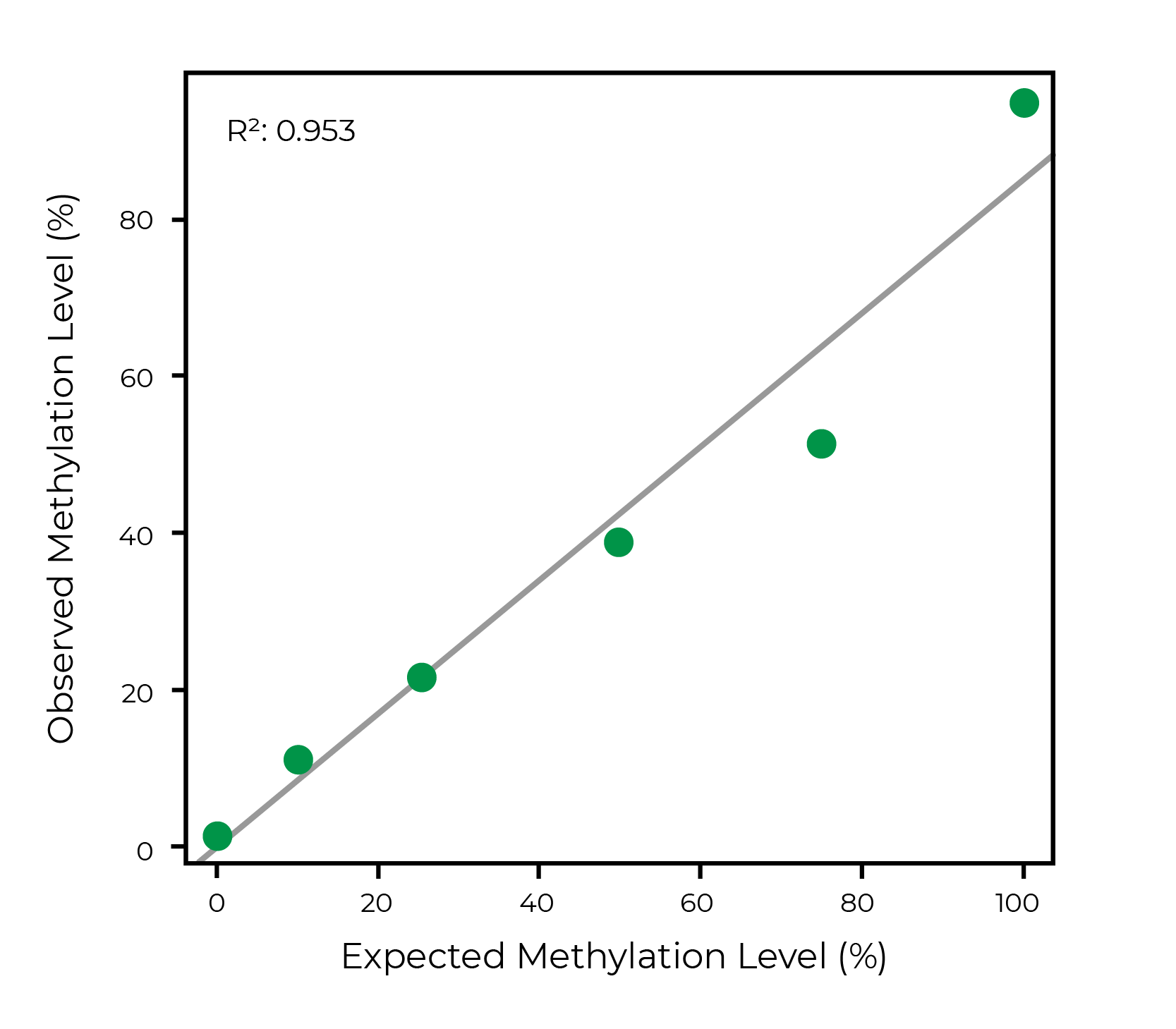
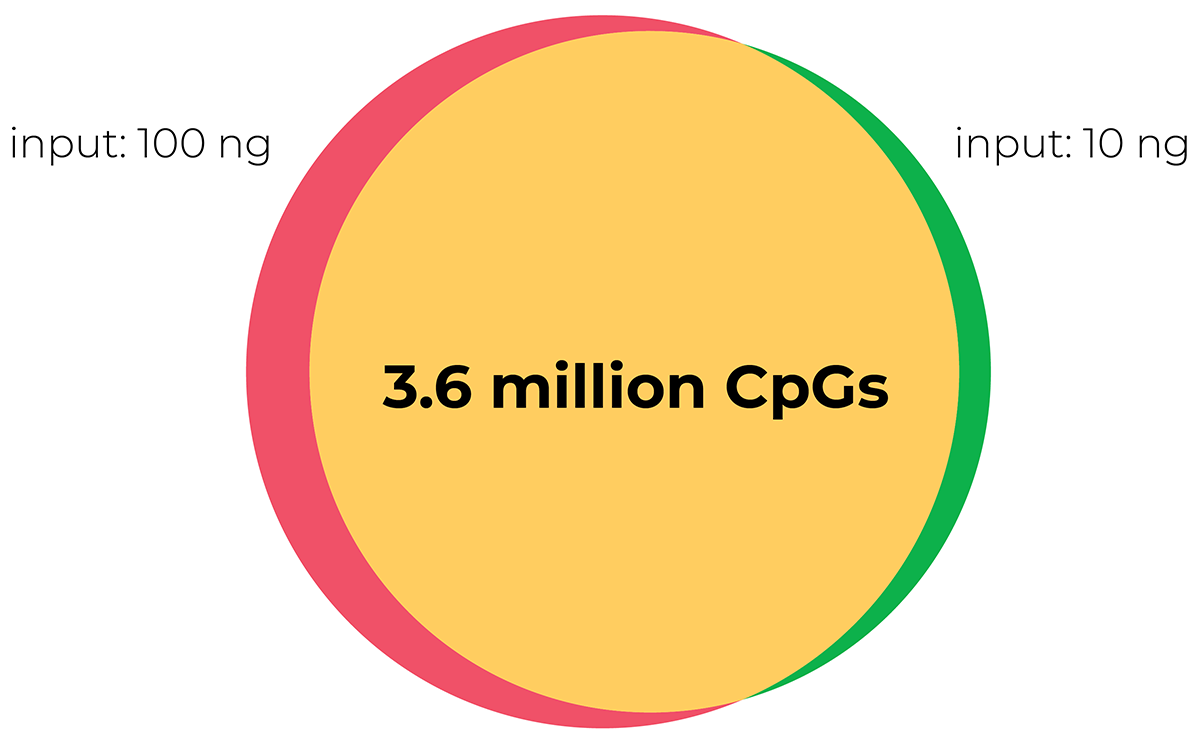
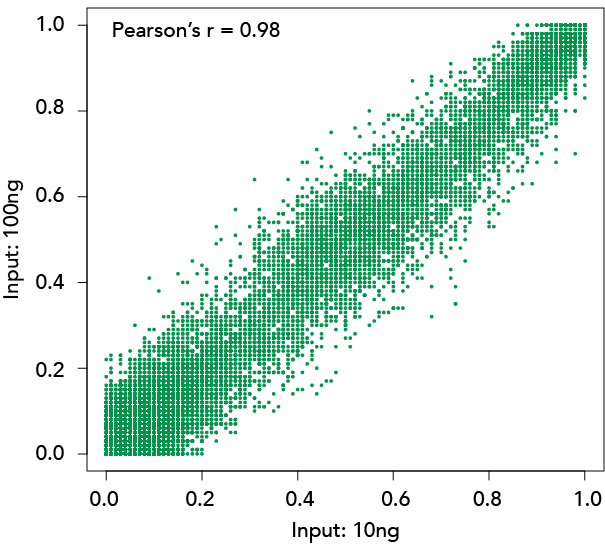
陆,Y。et al。(2020)重组恢复年轻的表观遗传信息和恢复视力。自然588年,124 - 129 (2020)
温德尔·p . et al。(2020)具有成本效益的方法DNA甲基化的检测甲基敏感的飞镖测序。《公共科学图书馆•综合》15 (6):e0233800。
https://doi.org/10.1371/journal.pone.0233800
研究人员研究表观遗传调控myonuclear利用rrb急性肥厚性刺激的反应。冯·沃尔顿f . et al。发现655 hypomethylated和84 hypermethylated网站从微分甲基化网站3000强启动子。途径分析了差动启动子区域甲基化显示强劲hypomethylation网站影响骨骼肌增长的监管机构,自噬和核糖体生物起源。
冯·沃尔顿f . et al . (2020) Myonuclear DNA Methylome急性肥厚性刺激的反应。表观遗传学。2020年4月,DOI: 10.1080 / 15592294.2020.1755581
Beetch m等人研究DNA甲基化模式的不同阶段诱发犬来帮助定义三阴乳腺癌原位(DCIS)发展。Methyl-Miniseq®利用发现68 128和96 150与差异甲基化基因启动子特定于原位乳腺管癌和浸润性导管癌。差异甲基化基因转录调控、细胞凋亡、信号转导和细胞迁移。
Beetch, M。、Harandi-Zadeh年代。、杨、t . et al . DNA甲基化景观的三阴性导管原位癌(DCIS)进展的入侵阶段犬乳腺癌。Sci代表2415 (2020)。https://doi.org/10.1038/s41598 - 020 - 59260 - 4
小约翰,b . et al .产前运输应激的影响差异DNA甲基化的生理控制行为和应激反应在乳儿婆罗门牛小腿,动物科学杂志》,98卷,问题1,2020年1月,skz368,https://doi.org/10.1093/jas/skz368
张问et al。(2019)基因突变的动态变化景观保护的核心套细胞淋巴瘤细胞的突变。前面。肿瘤防治杂志。9:568。doi: 10.3389 / fonc.2019.00568
冈萨雷斯,et al。(2019)。叶酸代谢影响植物免疫通过1 c蛋氨酸Synthase-directed全基因组DNA甲基化的提高。分子植物。doi: 10.1016 / j.molp.2019.04.013
培根,e . r . et al。(2019)。神经内分泌衰老之前准更年期,是由DNA甲基化。74年老化,神经生物学,213 - 224。doi: 10.1016 / j.neurobiolaging.2018.09.029
小约翰,b . p . et al。(2018)。产前交通压力改变基因组DNA甲基化在乳儿婆罗门牛小牛。动物科学杂志》上。doi: 10.1093 /雅/ sky350
Arechederra, m . et al。(2018)。基因甲基化的身体CpG岛预测高剂量的功能性致癌基因在肝癌。自然通讯,9 (1)。doi: 10.1038 / s41467 - 018 - 05550 - 5
Mishra, r . et al。(2018)。前列腺癌基质表观遗传改变驱动代谢和神经内分泌改变。中国投资,128 (10):4472 - 4484。doi: 10.1172 / JCI99397
考尔,g . et al。(2018)。可变剪接的helicase-like转录因子(Hltf):内含子retention-dependent活化的免疫耐受feto-maternal接口。PLOS ONE, 13 (7), e0200211。doi: 10.1371 / journal.pone.0200211
张,Na。et al。(2018)。肝段H19长非编码RNA的表达升高导致糖尿病高血糖。JCI洞察力。3 (10):e120304。doi: 10.1172 / jci.insight.120304
Ryu, t . et al。(2018)。继代的表观遗传景观适应海洋变暖。自然气候变化,8 (6),504 - 509。doi: 10.1038 / s41558 - 018 - 0159 - 0
约根森,伞齿轮等。DNA甲基化,通过DNMT1,发展的一个至关重要的角色,胃肠道平滑肌细胞和疾病。细胞死亡说474 (2018)。doi: 10.1038 / s41419 - 018 - 0495 - z
Verma d . et al。(2018)。全基因组DNA甲基化分析识别差异甲基化在冷漠银屑病表皮。皮肤病学研究学报,138 (5),1088 - 1093。doi: 10.1016 / j.jid.2017.11.036
张,z . et al .结构性DNMT3A-mediated新创DNA甲基化的基础。大自然554年,387 - 391 (2018)。doi: 10.1038 / nature25477
水域,s . a . et al。(2017)。DNA甲基化的景观有袋类动物x分子生物学与进化,35 (2),431 - 439。doi: 10.1093 / molbev / msx297
Yaish, m . w . et al。(2018)。全基因组DNA甲基化分析响应模型中盐度植物哈里发医生(Medicago truncatula)。BMC基因组学、19 (1)。doi: 10.1186 / s12864 - 018 - 4484 - 5
谢,k . et al。(2018)。表观遗传改变长寿监管机构,寿命降低,加剧导致的旧的父亲的后代小鼠体内病理。美国国家科学院院刊》上,115 (10),E2348-E2357。doi: 10.1073 / pnas.1707337115
Al-Harrasi, i . et al。(2018)。微分DNA甲基化和转录资料在枣椰树根暴露于盐度。PLOS ONE, 13 (1), e0191492。doi: 10.1371 / journal.pone.0191492
琼斯,c . m . et al。(2018)。全基因组DNA甲基化的描述浸润性鳞翅类害虫,棉铃虫Helicoverpa armigera。G3:基因| |遗传学、基因组g3.1112.2017。doi: 10.1534 / g3.117.1112
麦考密克h . et al。(2017)同基因的小鼠表现出性别差异在多个组织DNA甲基化模式。BMC基因组18,966年。https://doi.org/10.1186/s12864 - 017 - 4350 - x
巴赫,et al。(2017)。怀孕期间后果提供甲基捐助者的methylome后代从哺乳期和非哺乳奶牛。《公共科学图书馆•综合》,12 (12),e0189581。doi: 10.1371 / journal.pone.0189581
意图,m . et al。(2018)。选择性抑制CTCF绑定的iAs指导TET-mediated iAs-transformed 5-hydroxymethylation模式细胞重新编程。毒理学和药理学应用,338年,124 - 133。doi: 10.1016 / j.taap.2017.11.015
德沃金,m . et al .(2017)分析methylomes高地和低地柳枝稷(黍virgatum)使用MeDIP-seq和BS-seq生态型。BMC基因组18,851年。doi: 10.1186 / s12864 - 017 - 4218 - 0
埃里森,E.M. et al。(2017)单碱基决议的映射5-Hydroxymethylcytosine修改海马体的阿尔茨海默氏症。J摩尔63年>,185 - 197。doi: 10.1007 / s12031 - 017 - 0969 - y
Moghadam,香港et al。(2017)的早期生活应激影响Methylome和转录组的大西洋鲑鱼。5023年Sci代表7日。doi: 10.1038 / s41598 - 017 - 05222 - 2
梅森,A.G. et al。(2017) SMCHD1调节一组有限的常染色体上基因簇染色体。骨骼肌7日12。doi: 10.1186 / s13395 - 017 - 0129 - 7
Korkmaz F.T. et al。(2017)全基因组甲基化分析显示差异甲基化位点与年龄相关性增加牛相关成纤维细胞对有限合伙人的回应。BMC基因组18,405年。doi: 10.1186 / s12864 - 017 - 3796 - 1
Begue, g . et al。(2017)。DNA甲基化评估从人类缓慢,增大骨骼肌纤维。应用生理学学报,122 (4),952 - 967。doi: 10.1152 / japplphysiol.00867.2016
Nagymihaly, m . et al。(2017)。Ploidy-dependent共生细胞的表观基因组的变化与特定的基因表达模式。美国国家科学院院刊》上,114 (17),4543 - 4548。doi: 10.1073 / pnas.1704211114
意图,m . et al。(2017)全基因组DNA甲基化改变针对无机砷抑制CTCF链接绑定,DNMT表达和细胞转变。41474年Sci代表7日。https://doi.org/10.1038/srep41474
Mutoji, k . et al。(2016)。TSPAN8表达区别睾丸精原干细胞在青春期前的老鼠。生物的繁殖,95 (6),117 - 117。doi: 10.1095 / biolreprod.116.144220
多尔蒂,et al。(2016) CD4 + T细胞methylome导致一个明显的CD4 + T细胞在bovis-infected牛分枝杆菌转录签名。31014年Sci代表6日(2016年)。doi: 10.1038 / srep31014
钟,et al。(2017)二甲双胍改变DNA甲基化全基因组通过段H19 / SAHH轴。致癌基因36,2345 - 2354 (2017)。doi: 10.1038 / onc.2016.391
Trella, et al。(2016)。全基因组分析确定微分Leprel2启动子甲基化,Foxf1, Mmp25 Igfbp6, Peg12小鼠病变。骨科杂志》上的研究,35 (5),947 - 955。doi: 10.1002 / jor.23393
Crampton, m . et al。(2016)。分析Methylomes来自中美洲菜豆(菜豆l .)使用MeDIP-Seq和全基因组Bisulfite-Sequencing钠。在植物科学前沿,7。doi: 10.3389 / fpls.2016.00447
Barua, s . et al。(2016)。DNA甲基化碱基分辨率分析揭示了妊娠期补充叶酸影响老鼠的后代小脑的表观基因组。神经科学前沿,10。doi: 10.3389 / fnins.2016.00168
Steyaert, s . et al。(2016) epigenetically调控基因的全基因组搜索使用MethylCap-seq和RNA-seq斑胸草雀。20957年Sci代表6日。doi: 10.1038 / srep20957
周,j . et al。(2015)段H19 lncRNA宽改变基因组DNA甲基化调节S-adenosylhomocysteine水解酶。Nat Commun 10221。doi: 10.1038 / ncomms10221
Weyrich, a . et al。(2015)。父亲的代际表观遗传响应热暴露在野外雄性豚鼠。分子生态学,25 (8),1729 - 1740。doi: 10.1111 / mec.13494
Kozlenkov, a . et al。(2015)。大量的DNA甲基化差异的两个主要在人类大脑神经元亚型。核酸研究,44 (6),2593 - 2612。doi: 10.1093 / nar / gkv1304
研究的作者研究DNA甲基化变化4周后小鼠骨骼肌的经常锻炼。使用Zymo研究Methyl-MiniSeq平台,Kanzleiter等人发现3692不同启动子区域甲基化论文认定分布在2762。加上基因表达谱,综合分析显示361个基因中,DNA甲基化的变化与基因表达的变化。这些基因包括相关肌肉的生长和分化以及代谢调节。
Kanzleiter T, et al。(2015)运动训练改变dna甲基化模式相关的基因在小鼠肌肉的生长和分化。是杂志性金属底座。15,308 (10):E912-E920。doi: 10.1152
研究人员试图更好地理解表观遗传变化,发生在肝星状细胞(hsc)激活。格策等人刺激HSC激活在离体培养条件下通过改变。全基因组DNA甲基化模式被使用Methyl-MiniSeq服务从Zymo研究决定。数据显示大约400差异甲基化区域,根据大卫通路分析,包含在HSC激活基因富集过程相关。此外,locus-specific验证测试表明,DNA甲基化与基因表达变化的基因选择。研究人员继续表明,DNA甲基化的变化可能积极的结果,而不是被动的,脱甲基作用机制。
格策,S。et al。(2015)表观遗传改变在肝星状细胞活化。PLOS One。2015年6月12日。DOI: 10.1371 / journal.pone.0128745
研究人员调查了酒精如何影响所需的监管机制正常干细胞分化和胚胎发育。研究的作者利用全基因组DNA甲基化分析Zymo研究提供的服务来帮助描述表观遗传和基因表达变化的人类胚胎干细胞(为)接触少量的酒精(乙醇,EtOH)。全基因组分析显示,酒精暴露大量区域启动子甲基化引起的,特别是影响染色体片段的2,16日和18日在未分化和微分表达式为其与控制。这项研究的作者还发现,EtOH暴露影响关键通路参与正常的细胞功能,如新陈代谢和减少hESC多能性。
哈立德,O, et al。(2014)基因表达特征受饮酒导致的DNA methylomic放松管制在人类胚胎干细胞。干细胞研究。12 (3):791 - 806。
影响的调查人员di - (2-ethylhexyl)邻苯二甲酸酯(DEHP),一个无处不在的增塑剂和已知的内分泌干扰物,DNA甲基化改变成年雄性大鼠肾上腺的暴露在子宫内DEHP。使用Methyl-MiniSeq™服务从Zymo研究中,研究人员发现了972个不同的甲基化论文认定2183479 CpG网站调查的整个基因组。大多数差异甲基化论文认定被发现在CpG岛(40%)。有趣的是,第二个最高的频率差异甲基化论文认定被发现在岸边/货架区域(30%)。相比之下,没有包含大量不同启动子区域甲基化论文认定,这表明DEHP目标外遗传性区域调节组织的表达。不同甲基化的研究人员还发现了几个热点论文认定,恰逢地区差异表达基因。两个这样的热点在chr20p12与放松管制的相关基因的表达。这个区域包含基因免疫系统的本征函数的关键。研究结果支持了这样的观点,即典型的DEHP目前的环境可能会影响大量的表观基因组区域。
Martinez-Arguelles, DB &帕帕多普洛斯诉(2014)识别热点的DNA甲基化在成年男性肾上腺反应在子宫内暴露于无处不在的内分泌干扰物)增塑剂邻苯二甲酸二- (2-ethylhexyl)。内分泌学。1(1):124 - 33所示。
研究人员利用Zymo研究亚硫酸氢甲基MiniSeq和有针对性的测序服务识别和验证甲基化标记与结直肠癌(CRC)的非洲裔美国人。全基因组甲基化分析甲基MiniSeq透露355年13个基因差异甲基化CpG网站除了hypomethylated长点缀元素(线)CRC样本。此外,这些基因包含六个最高的50个论文认定度差异甲基化,然后进一步研究使用42和有针对性的酸性亚硫酸盐测序样品。更大的样本集和更有针对性的测序,作者能够验证四个基因,EID3, BMP3, GAS7, GPR75,小说在美国黑人患者CRC甲基化标记。
Ashktorab H, et al . (2014) DNA methylome分析识别小说甲基化基因在非裔美国人结直肠肿瘤患者。表观遗传学。9 (4):503 - 512
Methyl-MiniSeq服务是被作者用来确定DNA甲基化在基因表达稳定的胚胎干细胞细胞之间的过渡状态。Rex1多能标记的表达,被发现与甲基化表达的关键因素:Dnmt3b Tet1, Prdm14。Rex1-low Methyl-MiniSeq数据显示,细胞状态相比有较高的启动子甲基化在Rex1-high状态。差异甲基化发生在关键的推动者ESC Esrrb等监管机构,Tet1, Tcl2;这些结果与观察到的基因差异表达单一的分子RNA-FISH (smFISH)。此外,细胞DNA甲基转移酶淘汰赛或者处理DNA甲基化抑制剂Rex1-low有更少的细胞状态。这些发现表明,甲基化需要两个细胞之间的过渡状态及其维护。
歌手,Z, et al .(2014)动态异质性和DNA甲基化在胚胎干细胞。摩尔细胞。55 (2):319 - 331。doi: 10.1016 / j.molcel.2014.06.029。
研究人员检查口腔角质细胞的表观遗传修饰后eb病毒(EBV)感染扩大和精简版本的使用减少了表示酸性亚硫酸盐测序和生物信息学分析(Methyl-MiniSeq)。组发现EBV感染引起持久的DNA甲基化变化CpG岛,基因启动子,在身体相比未感染的细胞。此外,DNA甲基化的变化与损益影响基因的表达的子集。研究结果提供机械的见解如何EBV感染导致DNA甲基化变化通常观察到EBV-associated癌,暗示了潜在的未来epigenetics-based疗法。
Birdwell CE、et al .(2014)全基因组DNA甲基化是表观遗传不灭的角质形成细胞直接接触的eb病毒感染的结果。J性研究。pii: JVI.00972-14。
Methyl-MiniSeq (expanded-RRBS)服务从Zymo研究被用来研究干扰血液流动(维)epigenetically调节内皮细胞基因表达,导致内皮细胞来启动和维持斑块,导致动脉粥样硬化的发展。使用Methyl-MiniSeq平台,研究人员发现,部分颈动脉暴露于维表现出更高水平的hypermethylated CpG网站相比,部分颈动脉不暴露于维。方法还透露,治疗小鼠DNA甲基转移酶(DNMT)抑制剂5阿扎hypermethylated CpG网站的数量减少内皮部分暴露于维。这些发现表明DNMT维的机制诱导内皮细胞全基因组DNA甲基化变化。
邓恩J, et al . (2014) Flow-dependent后生DNA甲基化调节内皮细胞基因表达和动脉粥样硬化。中国投资。124 (7):3187 - 3199。doi: 10.1172 / JCI74792。
使用Methyl-MiniSeq服务,研究的作者调查BCL6致癌基因有助于弥漫型大b细胞淋巴瘤(DLBCL)的发展。研究者使用小鼠模型中Bcl6表达仅限于造血干/祖细胞(公司),发现Bcl6倾向发展中b细胞恶性的命运。生成的数据通过Methyl-MiniSeq服务帮助识别epigenetically重组公司的基因组DNA甲基化模式。拟议的BCL6“肇事逃逸”模型驱动的肿瘤发展提出了一个可能的原因为什么某些某些淋巴瘤患者的治疗策略失败而不是他人,表明潜在的治疗发展新途径。
绿色M, et al .(2014)瞬态表达式Bcl6足够的致癌作用和诱导成熟b细胞淋巴瘤。5:3904自然通讯。doi: 10.1038 / ncomms4904。
该研究的作者使用Zymo Methyl-MiniSeq服务识别基因组DNA甲基化模式的改变脑组织孕妇的饮食喂养的老鼠的后代相比,叶酸浓度高出10倍低叶酸控制。研究人员发现重大全球之间的DNA甲基化模式的改变两组老鼠幼崽。此外,研究显示显著水平的变化在两个CpG甲基胞核嘧啶以及CHG CHH CpG岛环境,基因启动子基因的身体,和一个非编码区碱基分辨率。在许多情况下,DNA甲基化改变的基因差异表达与定量实时PCR。研究结果表明,母体叶酸可能产生深远的影响在儿童和甲基化和发展模式对观察到的最近有实质性影响自闭症谱系障碍等疾病的增加。
Barua年代,et al。(2014)单碱基解决老鼠后代大脑methylome显示表观基因组的修改引起的妊娠叶酸。表观遗传学染色质,7 (1):3。doi: 10.1186 / 1756-8935-7-3。
的Methyl-MiniSeq®(扩大rrb)服务从Zymo研究被用来研究在小鼠全基因组DNA甲基化的变化成神经管细胞瘤肿瘤模型。作者发现癌症特异性区域相对较少,表现出DNA甲基化,这是不同于以前观察到的人类癌症。研究人员下一步调查全基因组DNA甲基化模式在其他类型的人类癌症的小鼠模型,其中包括乳腺癌伯基特淋巴瘤。他们发现不同的甲基化模式在鼠标模型比人类同行,这表明某些类型的癌症需要开发更好的动物模型用于研究相关的癌症诊断和治疗。
Diede SJ, et al。(2013)基本启动子CpG岛DNA甲基化差异人类癌症和癌症的转基因小鼠模型。表观遗传学。8 (12):1254 - 1260。
研究人员使用了Methyl-MiniSeq®服务从Zymo研究调查机制负责建立横纹肌肉瘤(RMS)肿瘤在儿童。横纹肌肉瘤是儿童骨骼肌癌症由于不完整的肌肉细胞分化形成。MyoD是一种细胞因子,通常驱动器正常骨骼肌细胞的分化,但不能在RMS细胞即使它表达到正常水平。在这份报告中,作者Methyl-MiniSeq使用®证明一个MyoD目标基因负责肌肉分化,JDP2,并不表示由于DNA甲基化在其启动子区域,和这种表观遗传沉默可能导致缺乏在横纹肌肉瘤细胞分化。
三联KL, et al。(2013)的比较基因组绑定MyoD人类肌原性的细胞在正常和横纹肌肉瘤标识区域和地方promyogenic转录因子抑制。摩尔细胞杂志。33 (4):773 - 84。
研究人员使用了Methyl-MiniSeq®服务从Zymo研究探讨DNA甲基化的多发性骨髓瘤的小鼠模型。与野生型控制相比,小鼠造血干/祖细胞ectopically MafB致癌基因表达进行了预设的特定表观遗传重编程激活血浆细胞癌变的命运。调查结果显示小说MafB-mediated“放手”多发性骨髓瘤表观遗传分子机制和相关的血浆肿瘤细胞起始,并为创新提供洞察力,有针对性的癌症治疗。
Vicente-Duenas C, et al。(2012)一种新的分子机制参与多发性骨髓瘤开发了针对MafB造血的祖细胞。EMBO j . 31(18): 3704 - 17所示。